

学会員論文紹介
近著論文の解説
論文サマリー
血管とリンパ管は、別々のネットワークを全身に張り巡らせ、それぞれ独自の機能を発揮します。両者は、最終合流地点である頸部の静脈角まで一切接続すること無く、各々が独立したネットワークを形成します。しかしながら、血管とリンパ管、特に静脈とリンパ管の特徴・構造を比べると、ほぼ見分けがつかないほど酷似しており、両者がお互いをどのように見分け、独立性を担保しているのかは、古くからの疑問として残されてきました。
本研究では、まず多発性肺嚢胞、腎がん、線維毛包種などを典型的症状とするBirt-Hogg-Dubé(BHD)症候群の原因遺伝子として知られるフォリクリン(Flcn)に関し、血管内皮細胞特異的欠損マウスを作成したところ、血管とリンパ管の異常吻合により胎生致死となる、という表現型を見出しました。この表現型は出生後のFlcn遺伝子欠失、つまり血管とリンパ管が完全に分離した後でも、再吻合してしまうことを見出しました(図1)。メカニズムとしては、静脈内皮細胞においてFlcnがリンパ管発生のマスター転写因子であるProx1の発現量を負に制御しており、この制御が破綻すると、『リンパ管もどき静脈内皮細胞』が生じ、静脈がリンパ管を接続すべき対象であると認識してしまうことを見出しました。さらには、Flcnによって核内移行が制御されるbHLH型転写因子Tfe3が、これまで報告の無かったProx1のエンハンサー領域に結合し、Prox1の発現を直接制御していることを見出しました(図2)。実際、遺伝子欠損マウスの重複交配により、生体でもこの経路がワークしていることが示されました。
本研究成果は血管・リンパ管という体内の2つの酷似する循環系が、なぜ一切交通することなく、独立したネットワークを形成するのかという、長年世界的に未解明とされている生物学的な疑問を解き明かしたという学術的重要性を持ちます。また、臨床的側面からは、がんの外科治療などの後遺症として起こるリンパ浮腫に対する治療への発展の可能性を秘めます。
著者コメント
まず、この場を借りて明記しておきたいこととして、この研究成果の第一発見者は約10年前、当時NIHでご一緒させていただいていた蓮見由紀子さん(co-first author)と馬場理也さん(co-last author)です。つまり『血管内皮でFlcnをノックアウトするとリンパ管に血液が充満する』という表現型の発見です。手軽に受理されそうな雑誌に投稿されようとしていたお二人を無理矢理説き伏せ、半ば強引に帰国後もマウスを解析させていただき現在に至ります。
留学当時を振りかえると、英会話がダメダメで(今でもですが)、もともとコミュ障気味の久保田にとって、「下ネタは古今東西万国共通」を座右の銘とし、それを日々実行して多国籍に交友関係を広げている馬場さんは、衝撃的でありミラクルでした。馬場さんにインスパイアされ、夜な夜なネットで英語下ネタ学習に励んでいたものの、結局自分で実行できなかったのは悔やまれます。現熊大IRCMS所長の須田先生に馬場さんを紹介したとき、まさか須田先生のラボにリクルートするとは思っておらず、諸々の事情を鑑みて慌てて止めに入ったのですが、時すでに遅し。ずっと後悔していましたが、須田先生からIRCMSで大車輪の活躍をされていると聞き、自責の念が和らぎつつあります。
本研究成果は、馬場さんだけではなく、その他多くの研究者の方々の多大なるご協力なしでは遂行できなかったものであり、この場を借りて心より感謝申し上げます。血管生物の先生方におかれましては今後ともご指導ご鞭撻のほどよろしくお願い申し上げます。
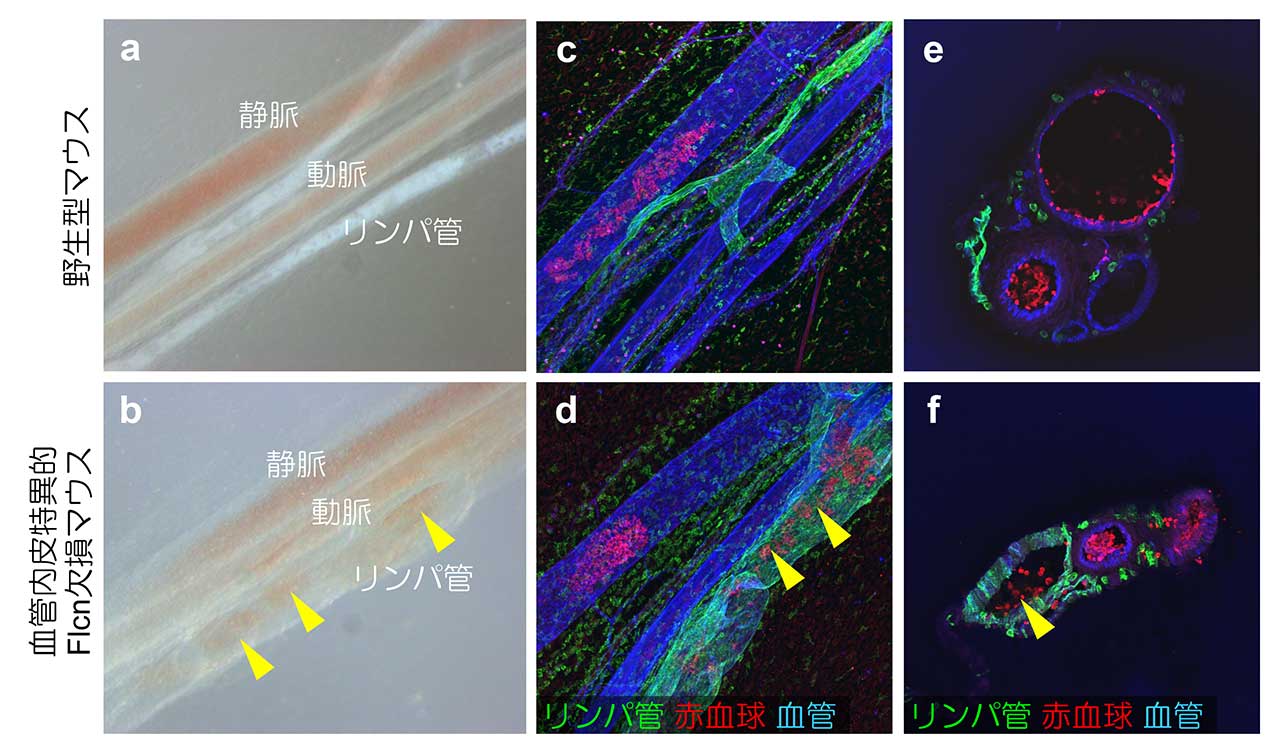
図1. 血管内皮特異的Flcn欠損マウスにおける血管―リンパ管異常吻合
野生型(a, c, e)および血管内皮特異的Flcn欠損マウス(b, d, f)における腸間膜画像。血管内皮特異的Flcn欠損マウスでは血管とリンパ管が異常吻合し、リンパ管内に赤血球が流れている。
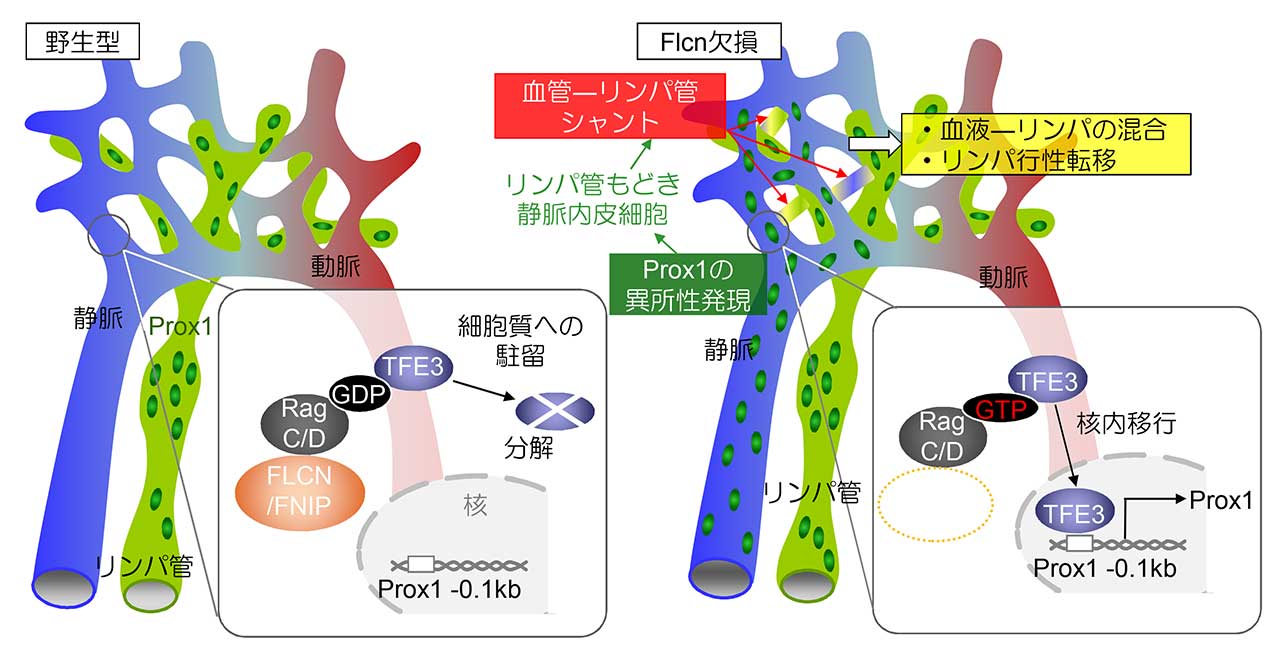
図2 . 血管とリンパ管の分離が維持される仕組み
通常、Flcnが転写因子TFE3を細胞質にとどめ、静脈におけるProx1の発現を抑えている。このためFlcnが欠失すると、TFE3が核内移行し、静脈でProx1が発現するようになり「リンパ管もどき静脈内皮細胞」が出現する。この細胞が原因となり、血管がリンパ管を接続すべき同志であると認識してしまう。つまり、Flcnは血管とリンパ管の可塑性を抑制する門番として働き、血管とリンパ管の分離を維持している。
-

後列左から5人目:田井育江(筆頭著者)6人目:久保田 -

左から2人目:馬場理也さん3人目:久保田